
15/12/2024
In the vast, unseen world that surrounds us, life thrives in forms so minuscule they often go unnoticed. Yet, these tiny inhabitants, particularly bacteria, are far from passive passengers in their environments. Instead, they exhibit an astonishing ability to navigate and respond to their surroundings with remarkable precision. This purposeful movement, known as taxis, is not merely random wandering; it is a sophisticated mechanism that allows bacteria to seek out favourable conditions, escape danger, and ultimately, survive and flourish. From sensing chemical cues to aligning with magnetic fields, the ways in which bacteria exhibit taxis are a testament to nature's intricate engineering at the microscopic scale.
Understanding Bacterial Motility: The Foundation of Taxis
Bacterial motility refers to the inherent capacity of bacteria to move independently, powered by their own metabolic energy. This fundamental ability is crucial for these microscopic organisms, enabling them to colonise new environments and discover vital resources for their survival. The movement of bacteria is influenced not only by the characteristics of the medium they inhabit but also by the various specialised appendages they employ for propulsion. Most rod-shaped bacteria possess this self-propelling power, a trait that has evolved in parallel among archaea, highlighting its evolutionary significance.
Diverse Mechanisms of Movement
Bacteria have developed a fascinating array of mechanisms to achieve locomotion, each adapted to different environments and purposes. These can broadly be categorised by the structures they utilise and the nature of their movement.
Flagella-Powered Locomotion
One of the most prominent and widely studied mechanisms for bacterial movement involves the use of flagella. These remarkable, hair-like filaments are responsible for two primary modes of locomotion: swimming and swarming.
- Swimming: This is the movement of individual bacterial cells within liquid environments. It's a highly efficient form of propulsion, allowing single cells to navigate freely through water or other fluids.
- Swarming: In contrast to swimming, swarming is a collective, multicellular 2D movement that occurs over solid or semi-solid surfaces. This coordinated translocation often requires the presence of surfactant substances, which help reduce friction and facilitate the rapid spread of bacterial populations. Swarming is a striking example of bacterial multicellularity and swarm behaviour, enabling rapid colonisation of new areas.
The flagellum itself is a helical, thin, and elongated appendage, anchored to the bacterial cell surface by a complex protein motor. This motor, situated within the cell membrane, causes the flagella to rotate at incredible speeds, ranging from 200 to 2000 revolutions per minute, depending on the bacterial species. A crucial component, the hook substructure, acts like a universal joint, connecting the motor to the flagellar filament, ensuring efficient power transmission. Prokaryotic flagella, unlike their eukaryotic counterparts which exhibit a bending motion, operate with a rotary movement. Bacterial flagella are typically powered by a proton gradient, converting electrochemical energy into mechanical work.
The arrangement and number of flagella on a bacterial cell can vary significantly, leading to different classifications of flagellation:
- Polar Flagellation: Flagella are present at one or both ends of the cell.
- Monotrichous: A single flagellum attached at one pole.
- Lophotrichous: A tuft of flagella located at one pole.
- Amphitrichous: Flagella present at both ends.
- Peritrichous Flagellation: Flagella are distributed widely across the entire cell surface.
The bacterial flagellum is a true protein nanomachine. Its basal body, a reversible motor, spans the bacterial cell envelope and consists of a central rod and several rings (L, P, MS, C rings in Gram-negative bacteria; only inner rings in Gram-positive). Mot proteins surrounding the inner rings facilitate ion translocation, providing the energy for rotation. Crucially, Fli proteins enable the flagellum to reverse its direction of rotation in response to specific environmental stimuli. The helical filament, composed of many copies of the protein flagellin, can rotate both clockwise (CW) and counterclockwise (CCW).
Pili-Mediated Movement: Twitching
Beyond flagella, bacteria also utilise hair-like appendages called pili (or fimbriae) for movement, particularly on solid surfaces. While the terms are often used interchangeably, pili are notably involved in a form of crawling motility known as twitching.
Twitching motility is characterised by the jerky, irregular motions of individual cells when observed under a microscope. This active movement is mediated by specialised structures called type IV pili. These pili extend from the cell's exterior, firmly attach to surrounding solid substrates, and then retract, effectively pulling the cell forward in a manner akin to a grappling hook. The cycles of polymerization (extension) and depolymerization (retraction) of type IV pili generate significant pulling forces, allowing bacteria to explore substrata and find suitable sites for growth, crucially aiding in the formation of bacterial biofilms. Pili also serve other vital functions, including cell-cell interactions, surface sensing, and even DNA uptake.
Other Modes of Surface Translocation
Not all bacterial movement relies on flagella or pili. Other fascinating mechanisms enable translocation across solid surfaces:
- Gliding Motility: This is a flagella- and pili-independent form of movement, allowing microorganisms to travel along the surface of low aqueous films. The precise mechanisms are still being elucidated, but it involves diverse motor complexes, such as the focal adhesion complexes seen in bacteria like Myxococcus. Gliding speeds vary between organisms, and direction reversal appears to be regulated by an internal clock.
- Sliding: Unlike twitching and gliding, which are active movements driven by individual cells, sliding is a passive movement. It relies on the collective motive force generated by the cell community itself, primarily due to the expansive forces caused by cell growth within the colony, especially in the presence of surfactants that reduce friction between cells and the surface.
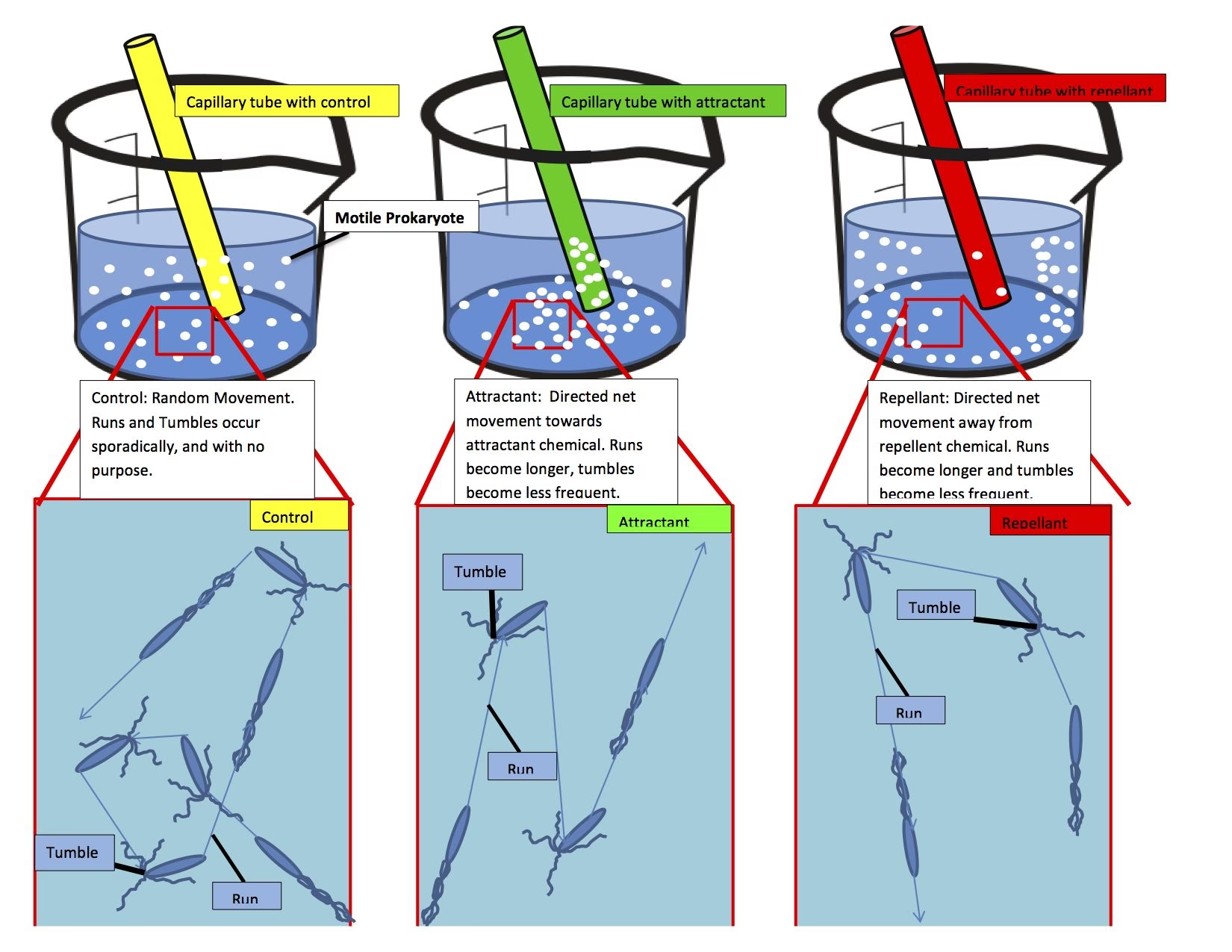
The general movement of a bacterium can often be described as a "random walk," characterised by alternating "run" and "tumble" phases. In a uniform environment, this results in relatively straight swims interrupted by random tumbles that reorient the bacterium. However, when bacteria exhibit taxis, this random walk becomes "biased" – directed towards or away from specific environmental stimuli. This allows bacteria to reposition themselves strategically.
Key Types of Taxis
Different types of taxis are distinguished by the nature of the stimulus that controls the directed movement:
Chemotaxis: Responding to Chemical Gradients
Perhaps the most well-studied form of taxis, chemotaxis, involves bacteria directing their overall motion in response to the presence of chemical gradients. Bacteria like E. coli, while unable to choose a specific swimming direction or maintain a straight line for long, employ a clever strategy: they use temporal sensing to evaluate whether their situation is improving. If a bacterium senses it is moving towards an attractant (or away from a repellent), it will prolong its straight swimming phase before tumbling. Conversely, if it's moving in the "wrong" direction, it will tumble sooner, reorienting itself randomly until a more favourable direction is found. This "biased random walk" allows them to find the highest concentration of attractants, even detecting subtle differences.
In marine environments, chemotaxis is profoundly important, as it enables microbes to navigate towards nutrient hotspots, directly impacting nutrient uptake and the cycling of elements in the ocean. Even in dilute conditions, where signal and noise are similar, bacteria must contend with the intrinsic randomness of molecular binding and signal transduction to navigate complex, dynamic landscapes of nutrient patches.
Phototaxis: Seeking or Avoiding Light
Phototaxis is the locomotory movement of an organism towards or away from a stimulus of light. This is particularly advantageous for phototrophic organisms, such as cyanobacteria, which rely on light for photosynthesis, allowing them to orient themselves for optimal light reception. Movement towards increasing light intensity is termed positive phototaxis, while movement away is negative.
Two forms of positive phototaxis are observed in prokaryotes:
- Scotophobotaxis: Observed only under a microscope, this occurs when a bacterium accidentally swims out of an illuminated area. Entering darkness signals the cell to reverse flagellar rotation and re-enter the light. It's an "escape from darkness" rather than direct light seeking.
- True Phototaxis: This is a genuinely directed movement up a light gradient, analogous to positive chemotaxis but with light as the attractant.
Photoreceptor proteins, such as bacteriorhodopsin and bacteriophytochromes, are light-sensitive proteins involved in sensing and responding to light. Halophilic archaea, like Halobacterium salinarum, utilise sensory rhodopsins (SRs) for phototaxis. These SRs trigger signalling cascades that ultimately influence flagellar rotation, allowing the cell to adjust its movement based on light intensity and quality (e.g., orange light as an attractant, blue or UV light as a repellent).
Magnetotaxis: Aligning with Earth's Fields
A truly unique form of taxis is magnetotaxis, where magnetotactic bacteria align their movement along the magnetic field lines of Earth. These bacteria possess specialised biomineralised organelles called magnetosomes, which contain magnetic crystals. This alignment is believed to assist these organisms in reaching regions of optimal oxygen concentration. It's important to note that unlike animal magnetoreception, which involves a stimulus-response mechanism, magnetotaxis in bacteria is due to fixed magnets within the cells, forcing alignment much like a compass needle, even in dead cells.
Escape Response: Rapid Retreat from Danger
An escape response is a rapid, negative taxis, allowing bacteria to back away from stimuli that could be harmful or lethal. This is fundamentally different from navigation or exploration, as it demands extremely rapid response times. Such reactions are not strictly oriented but commonly involve backward movement. Action potential-like phenomena have been observed in bacterial biofilms and in single cells, enabling these quick, phobic or emergency responses. For instance, Halobacterium salinarium exhibits a photophobic response involving a 180° reversal of swimming direction, triggered by changes in membrane potential.
Other Forms of Taxis
Beyond these primary types, bacteria exhibit other specific forms of taxis:
- Aerotaxis: The response of an organism to variations in oxygen concentration, predominantly observed in aerobic bacteria.
- Energy Taxis: This involves the orientation of bacteria towards conditions that optimise their metabolic activity. Unlike chemotaxis, which responds to extracellular compounds, energy taxis is driven by intracellular stimuli, such as the proton motive force, allowing bacteria to seek energetically favourable environments.
The Peculiar Physics of Microscopic Movement
Understanding how bacteria move requires delving into the unique physics of the microscopic world, a realm far removed from our everyday experiences. The incessant jiggling motion of pollen in water, first observed by Robert Brown in 1828, remained an enigma until Albert Einstein's work in 1905, which explained this "Brownian motion" as the result of molecular collisions.
A key concept in understanding movement at this scale is the Reynolds number (Re), a dimensionless ratio that quantifies the relative importance of inertia versus viscosity for fluid motion. For macroscopic swimmers like fish, inertia is dominant (Re = 100), allowing them to coast between propulsive events. However, for microscale swimmers like bacteria, viscosity overwhelmingly dominates (Re = 10-4). This means that for a bacterium, the fluid acts like treacle or thick syrup, and any inertial coasting effect is negligible, lasting mere microseconds and covering miniscule distances. As famously discussed by E. M. Purcell, this high viscous damping implies that only forces exerted in the present moment contribute to a microscale body's propulsion. Therefore, a constant energy conversion method is essential for bacteria to move.
Purcell's "Scallop Theorem" beautifully illustrates this principle. He considered a hypothetical scallop consisting of two rigid pieces connected by a hinge. He concluded that no matter how the scallop periodically opens and closes its hinge, it will always return to its exact starting point at the end of the cycle. This is because at low Reynolds numbers, reversing the direction of mechanical motion simply reverses all velocities in the system, preventing net displacement over a periodic cycle. This theorem highlights why bacteria cannot simply "flap" their way through water like larger organisms. Instead, they must employ non-reciprocal motions, such as the continuous rotation of their flagella, to achieve effective propulsion.
Comparative Overview of Bacterial Motility
To better understand the distinct features of bacterial movement, let's compare their various mechanisms:
| Motility Type | Primary Appendage/Mechanism | Environment | Nature of Movement | Key Characteristic |
|---|---|---|---|---|
| Swimming | Flagella (Rotation) | Liquid Environments | Active (Individual Cell) | Individual cell propulsion; run-and-tumble pattern. |
| Swarming | Flagella (Rotation) | Solid/Semi-Solid Surfaces (2D) | Active (Multicellular) | Coordinated, rapid population translocation; requires surfactants. |
| Twitching | Type IV Pili (Extension/Retraction) | Solid Surfaces | Active (Individual Cell) | Jerky, grappling-hook-like movement; crucial for biofilm initiation. |
| Gliding | Diverse Motor Complexes (e.g., Focal Adhesion) | Solid Surfaces (Low Aqueous Films) | Active (Individual Cell) | Flagella/pili independent; smooth, continuous movement. |
| Sliding | Cell Growth & Community Forces | Solid Surfaces | Passive (Community) | Relies on expansive forces from colony growth; surfactants reduce friction. |
Frequently Asked Questions About Bacterial Taxis
- What is the fundamental difference between swimming and swarming?
- Swimming involves individual bacterial cells moving through liquid environments, typically using flagella. Swarming, conversely, is a coordinated, multicellular movement of a bacterial population across solid or semi-solid surfaces, also powered by flagella but often involving cell elongation and requiring surfactants.
- How do bacteria "know" where to go in chemotaxis?
- Bacteria don't "know" in the human sense. Instead, they use a process called temporal sensing. They constantly compare the current concentration of a chemical (attractant or repellent) to what they experienced a moment ago. If conditions are improving, they continue swimming straight for longer; if conditions are worsening, they tumble and reorient more frequently, effectively biasing their random walk towards favourable gradients.
- Can bacteria swim backwards?
- Yes, some bacteria can. While many, like E. coli, primarily achieve reorientation through tumbling (random reorientation), others, such as Vibrio alginolyticus, can perform distinct forward and reverse swimming steps by reversing their flagellar motor. Some also exhibit "flicks" or turns by buckling for reorientation.
- Why is movement at the microscopic scale so different from ours?
- The key difference lies in the Reynolds number. For microscopic organisms like bacteria, the fluid environment is dominated by viscous forces, making it feel like thick syrup. Inertia, which allows larger objects to coast, is negligible. This means bacteria must continuously exert force to move, and simple reciprocal motions (like opening and closing a hinge, as in Purcell's Scallop Theorem) will not result in net displacement.
- What are magnetosomes?
- Magnetosomes are specialised, biomineralised organelles found within magnetotactic bacteria. They contain magnetic crystals, which allow these bacteria to align themselves passively with Earth's magnetic field lines, assisting them in navigating towards optimal oxygen concentrations in their environment.
Conclusion
The world of bacterial taxis is a compelling demonstration of nature's ingenuity. Far from being simple, passive cells, bacteria are sophisticated navigators, employing complex biological machinery – from the whirling flagella to the grappling-hook-like pili, and even internal magnetic compasses – to orchestrate their movements. Their ability to sense and respond to subtle environmental cues, whether chemical gradients, light intensity, or magnetic fields, is fundamental to their survival, allowing them to thrive in diverse and often challenging habitats. This intricate dance of microscopic life not only underpins ecological processes but also continues to inspire scientific exploration into biohybrid microswimmers, blending biology with human engineering to unlock new possibilities in miniature locomotion.
If you want to read more articles similar to Bacterial Taxis: Navigating the Microcosm, you can visit the Taxis category.